|
| Каталог статей |
|
ЛЕКЦИЯ № 4 ГЕНОТИП И ФЕНОТИПВесь предыдущий материал позволяет нам совершить переход от примитивной и ошибочной «мозаичной» схемы воплощения генотипа в фенотип, к реальной «волновой» схеме. Если бы фенотип был простой суммой отдельных признаков, то графически можно представить себе это в виде огромной мозаичной картины, где каждый отдельный признак соответствует одному паззлу мозаики, а свойства каждого паззла зашифрованы в одном из генов ДНК.
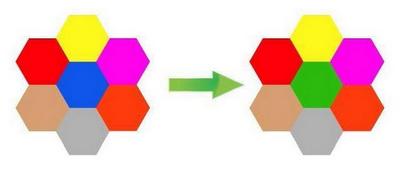
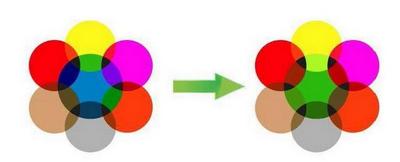
На рис. 10 изображен небольшой фрагмент мозаики, подразумеваем, что это часть масштабного живописного изображения. Произведем «мутацию», то есть поменяем синий паззл на зеленый, и увидим, что мутация в одном гене приводит к изменению цвета только одного участка, никак не влияя на соседние и лишь незначительно изменяя общий вид картины. А если взять схему ген-белок-клетка-признак, то следует обязательно учесть, что белок в клетке работает в тесном сотрудничестве с другими белками и изменение в его работе должно сказаться и на других белках и на самой клетке и далее на других клетках. Вместо дискретных стеклышек мозаики подойдут капли акварельной краски разных цветов (рис. 11).
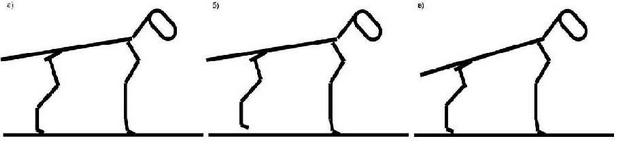
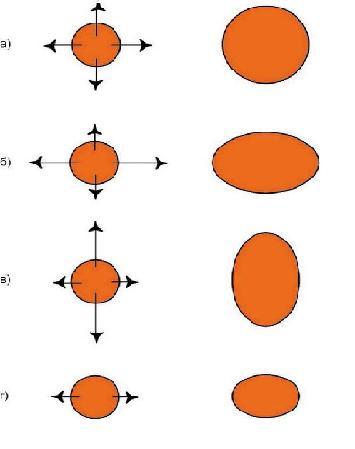
Первое что мы отметим: краски вступают друг с другом в контакт и в результате взаимодействия порождают совершенно новые цвета (признаки) за которыми не стоят никакие гены. Картинка сразу же становится богаче, чем в предыдущем примере, но и сложнее. Далее, мы видим, что при замене цвета акварельной капли, изменится не только один дискретный фрагмент, но и вся картинка. Если учесть, что в отличие от «признака», занимающего только одно конкретное место в фенотипе, многие белки могут находиться в клетках различных тканей, а, следовательно, и в совершенно разных компаниях других белков. А если еще вспомнить о том, что один ген может кодировать сразу несколько белков, то получится, что из-за одной единственной мутации волна изменений проходит по всей фенотипической картинке. Пример: у собак пигмент эумеланин бывает двух типов – черный и коричневый. Разница между ними даже не в химическом составе, а в способе полимеризации готового вещества. А теперь возьмем породы, в которых есть собаки в черном и коричневом вариантах одного окраса: лабрадоры, доберманы, ньюфаундленды, таксы, спаниели и пр. Опытные специалисты-породники подтвердят, что между черными и коричневыми собаками есть едва уловимые отличия. Вроде бы даже и не скажешь сразу, чем же они отличаются, но что-то такое есть, что даже заставляет в некоторых породах выставлять их в разных рингах. При этом иногда сотни лет идет общее разведение, так что прочие гены, находящиеся в той же 11-ой хромосоме уже должны были перемешаться, благодаря кроссинговеру (о К. будет ниже) и речь никак не может идти о сцепленном наследовании признаков. Причина исключительно в мутации в гене В. Казалось бы – такая чепуха, изменили всего лишь форму пигментных гранул с продолговатых на округлые, а вот какой резонанс во всем организме! Чтобы окончательно убедить читателей в несостоятельности мозаичной схемы и представлении о том, что за каждым признаком стоит если не ген, то группа генов, возьмем такой смешной пример как вера в существование генов, кодирующих углы конечностей и линии верха. Представим себе, что величины всех этих углов большими золотыми буквами четко прописаны каждый в своем гене и в зависимости от того, какая компания «угловых генов» соберется в одном генотипе, такие углы и будет иметь собака (рис. 12, а).
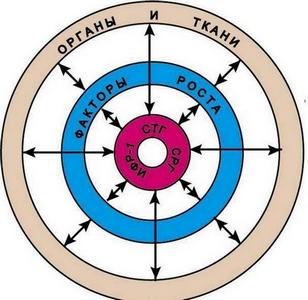
Но ведь кроме «генов углов» еще должны быть и гены, определяющие длины костей скелета! А теперь представим себе, что у собаки с означенными величинами «угловых», вдруг подкачал «ген бедра», и бедро оказалось коротковатым. Что, задние конечности собаки зависнут в воздухе (рис. 12, б)? Или может быть собака все же «упадет» на задние конечности (рис. 12, в)? Но ведь тут же возопит «ген линии верха»: «Господа, у меня указан совсем другой угол линии верха к горизонтали!». Генов углов гораздо больше, тем более, на их стороне гравитация, тянущая заднюю часть к грешной земле, так что они могут важно надуть щеки и сказать, что они эпистатичные гены, т.е. главные, доминирующие над генами другого локуса. И гену линии верха останется только горестно вздыхать: «Вы звери, господа! Что подумает какой-нибудь исследователь, заглянув в окуляр электронного микроскопа и увидев, что я обещаю одно, а на деле все совсем по-другому». Смешно. На самом деле, такой важный для собаководов признак как углы конечностей, это чисто виртуальный признак, зависящий исключительно от длин костей скелета и, частично, от сочленяющих их суставов. Но зато уж длины костей, безусловно, генетически зависимый признак. Так что, при оценке экстерьера собаки, при подборе племенных пар, нужен полный анализ анатомического строения. Ведь иногда правильные углы получаются благодаря удачно скомпенсированным недостаткам передних и задних конечностей. От умозрительных углов перейдем ко вполне конкретному и материальному глазу и рассмотрим принципы наследования его формы. Мы характеризуем глаз собаки следующими признаками: разрез (овальный и круглый), размеры (большие и маленькие), постав (косо и прямо поставленные). Давайте предположим, что за каждый признак отвечают специальные гены и сколько бы этих генов ни было, мы всех их вычислили. Ген это тоже вполне материальная вещь, реально существующий кусочек ДНК. И есть замечательный метод под названием – генная инженерия, когда вырезается определенный ген из ДНК одного животного и пересаживается в ДНК другого животного. В качестве подопытных кроликов возьмем шелти и пекинеса. У первых глаза узкие, миндалевидные, среднего размера, косо поставленные. У вторых - большие, круглые, выпуклые, поставлены прямо и широко. Можно взять оплодотворенные яйцеклетки чистопородного шелти и пекинеса и поменять им «глазные гены». Согласно «мозаичной» схеме у нас просто обязаны будут родиться самые обычные шелти, с типичными для породы узкими длинными головами, но…. на этих изящных головках будут красоваться крупные, круглые, выпуклые глаза пекинеса. А на широкой мордочке пекинеса, где-то в направлениях норд-норд-ост и норд-норд-вест расположатся две узенькие глазные щелочки. Представьте себе мысленно таких монстров, потому как рисовать мне их не хочется. Даже если вы не в состоянии объяснить – почему невозможно такое сочетание, то внутренний голос вам будет все равно твердить, что это невозможно, потому что невозможно никогда. И внутренний голос вас не обманывает. Давайте возьмем еще пример, более простой, на первый взгляд. Теперь мы произведем обмен только генами, контролирующими размер глаз, но зато сделаем это у йоркширского терьера и мастифа. Если мастифа с крошечными, как бусинки глазками еще худо-бедно представить себе можно, то что делать с йорком, ведь два мастифьих глаза больше всей головы крохи?! На самом деле нет, и не может быть никаких генов «глазных признаков». Расположение глаз, разрез, размер, постав зависят от глазницы. Где в черепе отверстие, какого размера и формы – такой глаз и будет. Глазница образуется на стыке 3 черепных костей и, значит, зависит от формы и размера этих костей. От конфигурации каждой кости черепа зависит и окончательная форма черепа. Так что наблюдается полная и абсолютная зависимость формы глаза от формы черепа собаки. Поэтому, если вы видите, что у собаки глаз не соответствует требованиям стандарта, но при этом вам кажется, что голова у собаки красивая, то это вам только кажется. Либо вам не хватает квалификации, чтобы увидеть недостатки в строении черепа, либо у вас глаз замылен, либо вам просто не дано видеть. Выходит, что форма глаза это вовсе не мелочь, на которую можно не обращать особого внимания при оценке собаки. Что же у нас получается в итоге? Расследуя механизмы наследования тех или иных признаков, мы не отказываемся совсем от генов, мы просто ищем - на что же на самом деле влияют гены и как это косвенно сказывается на фенотипе. В общих чертах схема формирования фенотипа была уже представлена в лекции по эмбриологии: каркас определяется hox-генами, грубые правки «фасада» производит некая группа важных генов, и мелкие штрихи добавляют все прочие гены. О роли hox-генов уже поговорили и выяснили, что они отвечают за внешний облик собаки: горизонтально расположенный корпус, четыре лапы, голова, шея, хвост. О том, как самый обычный ген, не связанный с анатомическим строением собаки, а отвечающий лишь за полимеризацию пигмента эумеланина, влияет на весь экстерьер особи, мы тоже рассмотрели. Осталась неисследованной лишь эта таинственная группа «очень важных генов». А это как раз и есть главные скульпторы в организме - факторы роста. Факторами роста (ФР) называют группу белковых молекул, которые стимулируют или подавляют размножение определенных типов клеток. Все вы, наверное, слышали о гормоне роста (соматотропный гормон, СТГ, соматотропин). Соматотропный гормон - вырабатываемый в передней доли гипофиза оказывает многообразное влияние на процессы обмена веществ, конечным результатом которых является усиление роста. Для того чтобы гипофиз приступил к синтезу СТГ, он должен получить команду из гипоталамуса – отдела головного мозга, координирующего работу многих органов. Происходит это посредством соматотропин-рилизинг-гормона (СРГ, соматолиберин). Произведенный гипофизом СТГ стимулирует гепатоциты печени вырабатывать инсулиноподобный фактор роста-1 (ИФР-1, соматомедин), а уж ИФР-1 в свою очередь регулирует работу гипоталамуса, таким образом, замыкая цепь. Опять мы сталкиваемся с очередной веселой троицей, повязанных друг с другом круговой порукой и заправляющих важнейшим для организма процессом – ростом, увеличением в размерах. СТГ, СРГ и ИФР-1– это три дирижера, три великих стратега, командующих общим процессом роста и решающим какого размера быть собаке. Гормоны поступают в кровь и вместе с током крови доходят до каждой клеточки организма, воздействуют на мембранные рецепторы и заставляют клетки делиться. Вроде бы все просто и понятно, но давайте все же присмотримся внимательнее к ходу этого процесса. Для этого мы слегка поиграем с пространственно-временным континуумом и усадим рядышком с новорожденным щенком его же самого, но уже выросшего и повзрослевшего. Сравниваем: кожный покров в процессе роста увеличивался преимущественно в плоскости и совсем немного в толщину; глазное яблоко практически идеальной сферической формы так и увеличивалось равномерно по всем направлениям; ухо новорожденного щенка любой породы напоминает по форме человеческий ноготь, далее растет только в 3-х направлениях - от черепа вверх или вниз, в зависимости от постава и в стороны; трубчатые кости растут преимущественно в длину и несколько меньше в ширину; кровеносные сосуды в длину и т.д. Разные ткани и органы растут по-разному, по своему индивидуальному сценарию. Гомоны дают команду «Расти!» и задают скорость роста, а многочисленные факторы роста на местах объясняют клеткам, как им расти, в каком направлении. Секретируются ФР самими же клетками, а вот воздействовать они могут как на свою клетку-родительницу, так и на другие клетки. Таким образом, факторы роста согласовывают процессы роста разных тканей. Опять посмотрим на нашего подопытного щенка – как росли его конечности? Рост трубчатых костей во весь период четко совпадал со скоростью роста прикрепленных к костям мышц. Не отстал и рост кожных покровов, покрывающих мышцы. Согласованность действий ФР ни в коем случае не является генетически зависимым процессом. В противном случае, если бы какой-то собаке достались гены, заставляющие активно расти трубчатые кости, а вот гены, активирующие рост мышц и кожи конечностей оказались слишком медленными, то зрелище получилось бы не для слабонервных – вылезшие наружу кости, разорванные мышцы и кожа. Взаимопонимание и взаимовлияние складывается в процессе «общения» факторов роста. Благодаря этому, даже если предприимчивый маленький кобелек исхитрится повязать крупную суку, никаких монстров для кунсткамеры от этого не родится. Это будут собаки некоего среднего размера, возможно недостаточно гармоничного строения, но уж точно не получится на огромном теле дога крошечной головы таксы. Ни естественным скрещиванием, ни с помощью генной инженерии нельзя присобачить цвергпинчеру овчарочьи уши, бладхаунду хвостик мопса, а к бедру чихуахуа прикрепить голень мастифа Условная схема роста может быть представлена в следующем виде:
Закольцованные гормоны и факторы роста стоят насмерть, не позволяя своим товарищам нарушить идеальную гармонию окружности. Если даже произойдет мутация в одном из ФР, то остальные факторы либо удержат своего модифицированного коллегу в прежних рамках, либо поддадутся его мощному напору и растянут или сузят свое кольцо. А когда происходит вязка партнеров значительно отличающихся размерами, то разномастные ФР, долей случая собравшиеся в одном организме, не станут тянуть одеяло каждый на себя, уродуя идеальную форму, а на самом деле – уродуя организм животного. Посовещавшись, они придут к консенсусу и выстроят среднеарифметическое кольцо. Даже если вы захотите далее поэкспериментировать с этим пометом и повяжете между собой сибсов, то вы не получите расщепления в потомстве на гигантов и миниатюру, как этого можно было бы ожидать исходя из законов Менделя. Это будут опять собаки среднего размера, хотя разброс в росте получится гораздо больший, чем при вязке двух собак такого же размера, но принадлежащих к устоявшейся породе. Но это еще не все функции ФР. Кроме регулировки процессов роста, эти белки обеспечивают согласованность действия иммунной, эндокринной и нервной систем. Нервная система – это собственно мозг и сеть нейронов, пронизывающих все тело. Нервная система координирует деятельность всех систем организма и обеспечивает возможность реализации психических процессов: мышление, память, эмоции. Эндокринная система состоит из ряда желез и является основной регулирующей системой, контролируя и интегрируя деятельность внутренних органов, посредством вырабатываемых гормонов. Иммунная система, состоящая из селезенки, костного мозга, лимфатических узлов, служит защитной системой и обеспечивает устойчивость организма. Вряд ли кто-то сомневался в том, что три эти системы связаны. Мы отлично знаем, что стресс может снизить иммунитет, а изменения в работе половых желез влияет на поведение, эмоции и даже на интеллект. Но информация о том, что связными работают именно факторы роста, позволяет нам понять - как каждый из более чем 200 тысяч белков в организме (и 20 тысяч их кодирующих генов), будь то иммунный белок или вообще пищеварительный фермент, могут косвенным образом повлиять на деление клеток, а, следовательно, и на фенотип. Также мы можем проследить связь между типом конституции и типом высшей нервной деятельности. Редкий собаковод не знаком с учением Павлова, и не отбарабанит как таблицу умножения 4 типа - 4 темперамента. Но задавались ли вы хоть раз вопросом: а почему именно так они связаны? И почему подобная зависимость наблюдается во всех породах собак, не смотря на заметные генетические различия? Да что там собаки, учение Павлова оказывается справедливым и для остальных видов млекопитающих, в том числе и человека. Если мы посмотрим через прицел факторов роста, то сможем объяснить связь: ФР, стимулирующие рост костей вширь сверх «сангвинической нормы», также оказывают тормозящее влияние на процессы в нервной системе. И наоборот – те ФР, что стимулируют рост скелетных костей больше в длину и меньше нормы в ширину, стимулируют скорость процессов в нейронах. Нельзя не восхититься в очередной раз великой мудрость природы, так удачно все устроившей. И это отнюдь не случайность. Более мощное и сильное животное не должно легко впадать в состояние гнева, чтобы не быть источником опасности для своих сородичей. Сухой и облегченной особи можно пробиться в жизни только за счет высокой скорости реакций, напора и агрессии. А слабым и нежным особям ни к чему ни сильное возбуждение, ни чрезмерное торможение, им будет легче выжить, если соплеменники как можно меньше станут их замечать. Мутации в ФР чаще всего являются породообразующими мутациями. Собака – прямой потомок волка и до того, как начался процесс образования разнообразных пород, все первобытные собаки были окрасом и экстерьером похожи на своего прародителя и на некоторых современных лаек. Как древнее человечество выводило новые породы собак, мы хоть и не видели своими глазами, тем не менее, можем описать с высокой степенью точности. Когда у собак происходили те или иные мутации, то мутантные особи отбирались и вязались друг с другом или схожими животными; носители нового признака шли в разведение далее, а неудачные особи отсеивались. Так постепенно новый признак закреплялся в ограниченном поголовье, которое с некоторой натяжкой можно назвать породой. И только в такой последовательности, а вовсе не как представляют себе некоторые собаководы: какой-то талантливый селекционер задумал получить собак с укороченной мордой, затем стал хитроумным способом подбирать пары обычных длинномордых собак и вдруг чудесным образом родилось желаемое. Или надоело человечеству до тошноты унылое однообразие в окрасах собак – все зонарные да зонарные, захотелось черных, рыжих, белых, коричнево-подпалых. Поднатужилось человечество и «вывело» новые окрасы. Даже сейчас некоторых собаководов пробирает дрожь восторга в случаях, когда в каком-то заморском питомнике местные «мичурины» вдруг «выводят» собак нового для породы окраса. На самом деле не нужно никаких сверхчеловеческих мыслительных способностей, чтобы повязать таксу со спаниелем, а потом выдавать потомков этих метисов кремового окраса за чистокровных такс. Аналогично и в других породах. Человек не может создавать гены или индуцировать нужные мутации в них. Мы можем только использовать накопившиеся за тысячелетия мутации и комбинировать их определенным образом. И хотя пород на сегодняшний день уже около 500, но породообразующих мутаций на самом деле не так уж много. К породообразующим мутациям относятся: 1) Мутации, влияющие на размеры – миниатюрность и гигантизм. На сегодняшний день известна одна из мутаций, благодаря которой мы имеем некоторые миниатюрные породы. Как и следовало ожидать – это мутация в стане «стратегов», а конкретно в ИФР–1. Как ни странно, этот же мутантный ген обнаружили и у ротвейлеров, которых даже самая изощренная логика не поможет приписать к той-породам. Если бы мы не познакомились ранее с круговой порукой факторов роста, то ничего бы не оставалось, как опять поминать нехорошими словами генетику. А так, мы можем наглядно убедиться в могуществе договора между всеми ФР, действительно, даже мутация не позволит уменьшить породу, если все остальные участники договора будут иметь иное мнение по этому вопросу. Факторы, влияющие на увеличение размеров собак пока неизвестны. Возможно, гигантизм собак, а также и некоторые формы миниатюрности связаны не только с мутациями, но и с разным количеством копий генов. О вариациях числа копий генов пока не упоминалось, а это, между прочим, еще одно интереснейшее явление, серьезным образом влияющее на формирование организма. Многие гены в ДНК встречаются не в единственном числе, а имеют дубли. Чем больше количество копий гена, тем выше экспрессия белка, тем значительнее влияние на фенотип. У человека целых пять генов СТГ, плюс альтернативный сплайсинг и благодаря этому в крови присутствуют сразу несколько изоформ гормона. Это облегчает и специализацию в воздействии на разные ткани и дает достаточно приличные вариации в росте у людей безо всяких там мутаций. 2) Мутации, влияющие на рост костей черепа. Выделяют три основных градации: Мезокефалия или среднеголовость - умеренно длинный и широкий череп. Для собаки это формы близкие к волчьему и лайкоидному черепу. Брахицефалия или короткоголовость, особая форма головы с относительно слабым развитием продольного диаметра и большим поперечником из-за преждевременного закрытия венечного (коронарного) шва. Долихоцефалия, или скафоцефалия - узкий череп, вытянутый спереди назад. Развивается из-за слишком раннего окостенения стреловидного (саггитального) шва. Есть так называемый закон Вирхова, который гласит, что при преждевременном заращении костного шва происходит компенсаторный рост костей параллельно к его оси. Но кроме этого у собак есть еще довольно-таки специфическая форма головы, распространенная у охотничьих пород – спаниелей, гончих, легавых. Возможно, такая форма вызвана преждевременным закрытием других швов, скорее всего боковых. Существует также характерная для всех той-пород округлая голова со слабо развитыми челюстями и это, скорее всего, следствие недостатка СТГ в организме, так как подобные изменения формы черепа наблюдаются в других породах, когда вдруг рождается щенок намного мельче, чем обычный представитель породы. Брахицефалия в кинологии традиционно связывается не сколько с широким черепом, столько с укороченной мордой. Но при закрытии коронарного шва морда должна укоротиться автоматически. Представьте себе череп лайки в виде латексной маски из магазина смешных ужасов. Коронарный шов проходит между лобной и теменной костью близко к линии ушей. Возьмем за края этого шва – это область скул, и растянем маску в ширину – длина черепной коробки уменьшится, ширина увеличится и на фронтальном срезе череп приобретет слегка округлую форму. Но также растянется в ширину и уменьшится в длину морда, хотя мы ее даже пальцем не коснулись. Заодно посмотрим и на то, как изменится форма глазницы – вместо овального косо расположенного отверстия получится круглое и прямо посаженное. Вот так одним легким движением мы трансформировали сложнейшую объемную конструкцию и изменили десятки характеристик, которые мы называем признаками. Но это еще не все, теперь сопоставим ширину черепа с шириной корпуса. Нельзя не заметить, что существует прямая зависимость одного от другого. Все брахицефальные собаки достаточно ширококостны и обладают объемной грудной клеткой. Мезоцефальные породы имеют нормостеническое телосложение, а у долихоцефалов вытянутое астеническое строение. Вот так-так – закрылся преждевременно один черепной шов и не только череп, но и всю собаку вдруг растянуло вширь или ввысь. Удивительно это будет до тех пор, пока мы будем оперировать сугубо медицинскими понятиями. С точки зрения молекулярной биологии заращение костного шва и компенсаторный рост костей параллельно к его оси означает изменения в факторах роста, задающих направление деления клеткам плоских костей черепа. Схематично можно изобразить в следующем виде.
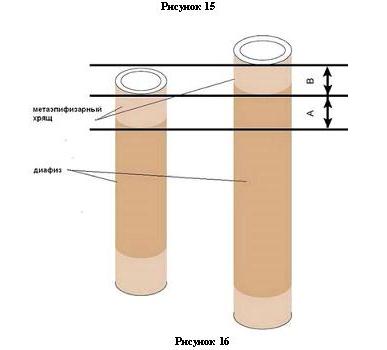
Есть исходная геометрическая фигура – круг. Для того чтобы в процессе роста круг сохранил свою форму, он должен расти равномерно по всем направлениям. Векторы силы, выполняющие в нашем случае роль факторов роста, должны быть равны (рис. 14, а). Теперь представим, что часть ФР, действующих в вертикальном направлении, вдруг поменяли пространственную ориентацию и стали стимулировать рост в горизонтальном направлении. Вместо круга получится овал, с горизонтальной большой осью и малой вертикальной – аналог брахицефального черепа (рис. 14,б). Изменим ориентацию горизонтальных ФР и получим аналог долихоцефального черепа (рис. 14,в). Пока нельзя сказать каким образом ФР плоских костей черепа влияют на рост остальных костей скелета. Возможно самым непосредственным – это один и тот же ФР, присутствующих во всех костях и естественно, мутация в нем приведет к одинаковым изменениям в росте костной ткани. А может через ту самую круговую пороку факторов роста, когда изменение в одном белке должны обязательно либо быть поддержаны остальными белками, либо запрещены. Независимо от того, каким путем согласуются процессы роста плоских и трубчатых костей, результат, как говорится, налицо: не бывает собак с широкой массивной головой и легким астеническим телом, как не бывает и мощных крепышей с утонченной длинной головой. Также невозможно к широкой и короткой черепной части приставить длинную морду, а к долихоцефальному черепу укороченные челюсти. И это прекрасно доказывается примерами межпородных вязок собак принадлежащих к разным типам. Как и в случае с межразмерными вязками – получается некий усредненный тип. Но все же можно найти несколько примеров, на первый взгляд, не согласующихся с выстроенной закономерностью. Пример первый: ротвейлер и боксер имеют приблизительно одинаковые размеры, головы у ротвейлеров заметно шире, чем у боксеров, но при этом и телосложение гораздо мощнее, что пока вписывается в предложенную модель. Морда у ротвейлера коротковата, что также пока не опровергает теорию. Вот только у ротвейлера отсутствует характерная для боксера вздернутость морды, вызванная специфическим искривлением лицевых костей. В этом нет ничего удивительного, это означает лишь, что у боксеров, бульдогов, пекинесов и пр. наблюдается дезориентация ФР и в лицевых костях, что приводит к их деформации в процессе роста. Пример второй: японский хин имеет ярко выраженный брахицефальный череп, а тело либо нормостеническое, либо вообще – астеническое. Но если мы посмотрим на ширину черепной коробки хина, то отметим, что голова у него вовсе не так уж и широка. У собаки с обычной головой и с таким же точно телосложением, ширина черепа была бы соразмерной. По-видимому, у хинов отсутствует компенсаторный рост черепа в ширину при прекращении роста в длину. ФР не меняют направление, а вообще прекращают работать (рис. 14, г). Пример третий: бультерьер на массивном, широком теле носит достаточно длинную голову. Однако назвать такую голову долихоцефальной никак невозможно. Голова бультерьера достаточно широка, а, кроме того, еще и наблюдается аномальное разрастание лицевых костей в высоту. Достаточно специфическое сочетание нескольких «потерянных в пространстве» ФР. Но все же не на столько они оказались потеряны, чтобы совсем не сообразить, что под такую массивную голову нужно обязательно соответствующее тело, что и было с успехом реализовано остальными ФР. 3) Хондродистрофия (ахондроплазия, коротконогость) – нарушение роста трубчатых костей конечностей. Вызвана мутацией в рецептора фактора роста фибробластов - FGFR3, который отвечает на воздействие факторов роста и контролирует производство протеина на поверхности клеток хрящевой и костной тканей. Есть много коротконогих пород собак и по ряду косвенных признаков можно предположить, что это разные мутации. По крайней мере, коротконогость у такс явно не та, что у терьеров или пекинесов. Данная аномалия встречается не только у собак, но и у многих домашних и диких животных, а также и у человека. Притом у людей где-то в 85% случаев это все новые мутации, что указывает на исключительную неустойчивость данного гена. Можно не иметь в роду хондродистрофиков и родить младенца с такой аномалией. Справедливо это и для собак. Не так уж уникальны случаи, когда от нормальных длинноногих собак рождался щенок с хондродистрофией. Конечно, если в облике такого щенка замечены черты одной из коротконогих пород, то виновата тут не мутация, а владелец разиня. Но если щенок яркий представитель своей породы и лишь длина лап не соответствует норме, то, вне всякого сомнения – это генетическая мутация. К счастью, эти мутации чаще всего доминантные, а значит можно уверенно полагать, что мутация произошла при формировании половой клетки или на ранней стадии эмбриогенеза, и родители не являются носителями неблагополучного гена в рецессиве. Но, на всякий случай, учитывая коварство и неустойчивость данного гена, следует обращать внимание – не родится ли далее по линиям обоих производителей еще раз такой же щенок. Это будет означать, что в породе возникла неприятная рецессивная мутация. 4) Бесшерстность – мутация в гене FOXI3 – фактор, контролирующий на эмбриональной стадии развитие клеток, из которых впоследствии формируются волосы и зубы. Так что FOXI3 одним выстрелом убивает сразу двух зайцев. Отсутствие зубов у голых собак это не следствие так называемого сцепленного наследования, когда два гена располагаются рядышком в хромосоме, а результат одной-единственной мутации в одном гене. И надо сказать спасибо, что эта мутация затрагивает лишь тот пул стволовых клеток, который превращается в зубы и шерсть. Да и то лишь у гетерозиготных особей. У гомозигот по мутантному аллелю нарушения эмбриогенеза настолько серьезные, что зародыши погибают. Исходя из этого, можно посоветовать тем заводчикам, которые все еще настойчиво пытаются «вывести» полнозубых голых китайских и мексиканских собак, направить энергию на решение реальных проблем. В крайнем случае, если уж так сильно хочется зубов, можно позаимствовать ген бесшерстности у американских голых терьеров. Это совершенно другая мутация, влияющая на какой-то другой белок, способный убить волосяной фолликул, но только лишь фолликул, а не другие клетки. Аналогичная мутация есть и у кошек – голые коты имеют полную зубную формулу. 5) Переразвитый кожный покров. Встречаются у бассетов, бладхаундов, шарпеев и др. По бассетам и бладхаундам пока сведений нет. Известна только причина, украшающая складками шарпеев - мутация в гене HAS2 (гиалуронат-синтаза). Гиалуронат-синтаза – белок клеточной мембраны, синтезирует гиалуроновую кислоту (ГК) – важнейший и интереснейший биополимер, входящий в состав многих тканей. В коже ГК принимает активное участие в регенерации ткани и стимулирует деление клеток кожного покрова. Будучи основой внеклеточного матрикса ГК поддерживает архитектуру тканей. Входит в состав стекловидного тела (глаз). Создает оболочку хрящевых клеток – хондроцитов. Главный компонент синовиальной жидкости суставов. Стоит перечитать список и сразу становится понятно, что муциноз кожи – это не сопутствующее заболевание шарпеев, а собственно то, что делает их шарпеями. Муциноз -отложение в коже муцина – желеподобного вещества, состоящего преимущественно из гиалуроновой кислоты. Избыток ГК как раз и создает складчатую кожу – породную особенность шарпеев. Также становится понятным, почему шарпеи склонны к отекам суставов и морды. И даже склонность к различным глазным заболеваниям: глаукома, смещение хрусталика, дисплазия сетчатки. HAS2 не двух зайцев подстрелил, а целое стадо. В свете этих данных борьба за чистоту породы от перечисленных заболеваний превращается в фарс. 6) Форма хвоста. Хвосты у собак демонстрируют богатое разнообразие форм: прямые(поленом или прутом); изогнутые (саблевидные, серповидные); с крючком на конце; кольцом, двойным кольцом, куцехвостость. Куцехвостость – уменьшение, искривление хвостовых позвонков, скорее всего связана с архитектурными генами hox12 и hox13, работающими в той части эмбриона, которая в дальнейшем станет хвостом. Это могут быть мутации и в самих этих генах, и в регулирующих их последовательностях. Остальные формы хвостов не связаны с изменением строения позвонков. Даже у собак, имеющих хвост кольцом, позвонки абсолютно нормальны. Изогнутость хвост приобретает из-за генетически обусловленных изменений в строении связочного аппарата хвостового отдела позвоночника. Связки плотно спаяны с телами позвонков и их функция состоит в контроле диапазона, в котором разрешено сгибаться позвоночному столбу. Следует отличать индивидуальную особенность некоторых собак – слишком высоко нести хвост, причиной тому излишний тонус хвостовых мышц. 7) Форма ушей. Как бы ни странно это прозвучало, но уши у собак отличаются только по длине и плотности хрящевого полотна. Вне всякого сомнения – это признаки, напрямую зависящие от каких-то факторов роста. Постав ушей, так же как и глаз, зависит исключительно от формы черепа. Где слуховое отверстие в черепе – там и окажется ухо. Будет ли ухо стоячим или висячим зависит в первую очередь от постава. Слишком низко, намного ниже линии лба, посаженное ухо всегда будет висячим. Ему физически просто некуда подняться вверх, мешает череп. Уши, расположенные на линии лба или немного ниже будут стоячими или висячими в зависимости от размера уха и качества ушного хряща. Небольшое, высоко посаженное, но тонкое и мягкое ухо фокстерьера повиснет на основании хрящей. Имеющие достаточную плотность хряща, уши дога и добермана упадут исключительно по причине излишней длины. Если купировать уши щенкам этих пород, убрав излишек массы, то они запросто превратятся в стоячие. Остался еще один показатель – форма уха. Уши у собак бывают треугольные и закругленные, но форма зависит лишь от длины уха, а не от дополнительных факторов. Относительно короткое ухо всегда будет треугольным, по мере увеличения относительного размера уха, треугольник начинает терять симметричность и плавно переходит в закругленную, лопухообразную форму. Не бывает коротких лопухообразных ушек и не бывает слишком длинных заостренных треугольных ушей. Треугольники ушей отличаются между собой – равносторонние, равнобедренные. Форма треугольника зависит от ширины основания и высоты. Высоту определяют факторы роста, заставляющие клетки хрящевой ткани делиться с заданной интенсивностью, а вот ширина уха зависит от морфологических изменений височной кости (опять таки под воздействием ФР), на которой располагается ухо. Так называемое, полукруглое ухо чау-чау на самом деле имеет треугольную форму, просто при таком соотношении высоты треугольника и ширины его основания, ребра располагаются под острыми углами, и получается фигура похожая на полукруг. Стоит какому-то чау отрастить себе ушки подлиннее, как сразу же они превращаются в обычные треугольники. Требование стандарта к форме ушей, по сути своей является требованием к длине уха. Просто термин «излишне длинные» можно истолковать как угодно, а вот указание формы уха получается более надежным ограничителем злоупотреблений. 8) Тип шерстного покрова. У собак наблюдается великое разнообразие и по длине шерсти, и по жесткости, прямые и кудрявые, наличие или отсутствие подшерстка, характер оброслости по телу. На сегодняшний день известны три гена — RSPO2, FGF5 и KRT71, мутации в которых влияют на рост шерсти. От гена RSPO2 зависит, будет ли собака брудастой, вырастут ли у нее «усы» и «брови»; FGF5 отвечает за длину шерсти; KRT71 — за курчавость. Разнообразные комбинации их определяют около 95% типов собачьей шерсти. Белок R-spondin 2 (RSPO2) регулирует формирование органов энтодермального происхождения: кишечник, легкие, трахеи, а также и волосяные фолликулы. К счастью, мутация произошла не в кодирующем экзоне, а в интроне и собаки получили лишь бородку и усы, благополучно избежав серьезнейших пороков развития внутренних органов, что неизбежно происходит при более серьезных мутациях в RSPO2. Белок FGF5 еще один фактор роста фибробластов. Мутация в этом гене продляет фазу роста волос (анаген). Но не только на рост шерсти влияет данный протеин, он обнаруживается в различных тканях во время эмбрионального развития. Потому в тех породах, где существуют длинношерстные и короткошерстные разновидности неизбежны некоторые отличия в экстерьере между ними. Экспертам и заводчикам следует это знать, не делить разновидности на «главную» и «второстепенную», «более породную» и «метисов», не требовать и не пытаться достигнуть полной идентичности собак с разными типами шерсти. Любопытно, что афганские борзые, обладающие особенно длинной шерстью, не имеют мутации в FGF5. Так что нас ждет обнаружение еще одного гена, влияющего на рост шерсти. Кудрявая шерсть у собак связана с изменением структуры кератиновых волокон, из которых состоят волосы. 9) Ридж. Специфический узор на спине у родезийских и тайских риджбеков получается благодаря дублированию длинного участка ДНК, на котором расположились 4 гена: FGF3, FGF4, FGF19, ORAOV1. Эти белки из семейства ФР регулируют в эмбриогенезе миграцию клеток нервного гребня. Также все эти белки связаны с канцерогенезом и относятся к группе онкогенов. Мы уже знаем, что увеличение копий генов усиливает экспрессию белка, в данном случае это приводит к нарушению развития эпидермиса вдоль хорды. При незначительных нарушениях меняется лишь пространственная ориентация волосяных фолликулов и шерсть начинает расти в нестандартном направлении. Между прочим, подобное встречается и у собак других пород, но в гораздо менее выраженной форме – маленькие в 1-3 см. риджи, и это не связано с вышеуказанной мутацией, скорее всего это мелкое нарушение в передвижении эктодермальных клеток в эмбриональном периоде. При более серьезных отклонениях происходит хаотичное и непредсказуемое перемещение клеток различных тканей, что вызывает крайне неприятную аномалию - дермоидный синус. Дермосинус представляет собой канал в коже, иногда доходящий до позвоночного столба. В полости дермосинуса встречаются волосы, жировая ткань, чешуйки эпидермиса. Доказано, что дермосинус не просто сопутствует риджу, но и частота его появления связана с экспрессией генов. У гомозиготных по вышеуказанной мутации особей вероятность образования синуса гораздо выше, чем у гетерозиготных. Дермоидный синус встречается и у собак без риджа, но в других породах это исключительно редкое явление. Удивляться этому не нужно – мало ли какие причины заставили группу стволовых клеток нарушить заданный маршрут и потеряться в дебрях организма. Бывают кисты и из других зародышевых листков, и даже сложные, состоящие из элементов всех трех видов экто-, эндо- и мезодермы, которые также никак не связаны с этой мутацией. 10) Окрас. Также является породообразующим признаком. Есть породы, особенно в группе гончих, которые отличаются одна от другой практически только одним окрасом. Мутаций, влияющих на окрас очень много, рассматривать их следует только в комплексе, потому нет возможности в рамках лекции подробно разобрать все эти гены и белки. Перечислю в общих чертах. Среди окрасообразующих, есть мутации влияющие на процесс миграции стволовых клеток предшественников меланоцитов; есть мутации в мембранных белках, в том числе и в рецепторах; есть мутации в белках воздействующих на рецепторы (лигандах); в транспортных белках, осуществляющих перенос как готовых пигментных гранул, так и компонентов для их изготовления; в белке полимеризаторе; в сигнальных белках. Вот, собственно, и все. На первый взгляд совсем не много, но если комбинировать все эти мутации в разных сочетаниях, то этого вполне хватит на создание нескольких сотен достаточно оригинальных пород. Лишь несколько породообразующих признаков являются следствием мутаций связанных с нарушениями эмбрионального развития. Остальные прямо или косвенно связаны с факторами роста. Даже мутация в HAS2. Гиалуронат-синтаза сама хоть и не относится к ФР, но управляющий сигнал получает непосредственно от факторов роста TGF-b и PDGF. Ну и, поскольку уж речь зашла о факторах роста, то давайте попробуем порассуждать о том, как ФР могут повлиять на появление такой неприятной аномалии как дисплазия тазобедренного сустава (ДТС). О дисплазии на сегодняшний день написано несметное количество статей, проведено множество исследований. Известные кинологи и ветеринарные медики сходятся во мнении, что причин для развития ДТС может быть несколько: генетические мутации; гормональные нарушения; недоразвитые сухожилия и слабая мускулатура таза; прямой постав бедра; растянутый формат; высокая скорость роста и избыточный вес; несбалансированное питание; чрезмерные физические нагрузки; травмы; воспалительные заболевания суставов и пр. Трудно с этим не согласиться. Мутации в белках, связанных с кальциевым обменом в организме запросто могут привести к дисплазии. Гормоны, регулирующие кальциевый обмен тоже важные и нужные участники процесса. Любые механические воздействия на молодой не окостеневший сустав могут привести к разрушению хрящевой и костной тканей. Сустав методом фотосинтеза из воздуха не создашь, потому в миске щенка должен присутствовать полный набор «стройматериалов» для размножения клеток костной ткани. Все щенки рождаются со здоровыми суставами, и лишь к 12-18 месяцам развиваются отклонения от нормы, то есть изменение происходит в процессе роста, а кто заправляет всеми процессами роста в организме, мы уже знаем. Теперь обратимся к статистике по частоте ДТС в разных породах собак. К сожалению, нет возможности привести полный список, но он доступен всем пользователям интернета на сайте Ветеринарной Ортопедической Организации (OFA). Если вы просмотрите список от начала до конца, то согласитесь, что тяжелые ширококостные породы располагаются преимущественно в верхней части списка, к средине их становится все меньше, а в конце списка в основном обосновались, стройные и высоконогие породы. Позиции пород, на первый взгляд не вписывающихся в выявленную закономерность, можно объяснить исходя из многообразия причин вызывающих ДТС, например, три мелкие породы - английский бульдог, мопс и французский бульдог, не смотря на свой небольшой вес, относятся к «широким» брахицефальным породам, да еще и имеют практически вертикальное бедро, при котором головка бедра просто не может правильно расположиться в вертлужной впадине. Но есть в этом перечне и труднообъяснимые пункты - достаточно высокий процент дисплазии у всех сеттеров. Возможно, в этих породах решающими являются другие факторы из представленного. Даже в обнаруженной закономерности есть определенные нестыковки, почему-то у намного более крупных и тяжелых немецких догов процент нарушений гораздо ниже, чем у золотистых ретриверов или ротвейлеров. Однако этому можно найти объяснение. Давайте рассмотрим строение бедренной кости. Эта кость, как и все трубчатые, кости скелета, состоит из двух частей - трубчатая цилиндрическая средняя часть (диафиз) (рис. 16), и утолщения на концах (эпифизы) (рис. 15). В щенячьем возрасте диафиз отделен от эпифиза метаэпифизарным хрящом и рост трубчатой кости в длину обеспечивается этой самой метаэпифизарной хрящевой пластинкой. Фактически, диафиз и эпифиз растут и развиваются отдельно друг от друга, лишь когда хрящевая пластинка истончается и исчезает, кость сливается в единое целое и рост ее в длину заканчивается. Диафиз и эпифиз имеют различное строение. В зависимости от положения костных пластинок различают - губчатую и компактную костную ткань. Трубчатая часть кости состоит преимущественно из компактного плотного вещества. Снаружи компактная костная ткань покрыта надкостницей, а внутренний слой переходит в губчатое вещество. Внутри кости находится полость, заполненная костным мозгом. Эпифиз состоит из большого количества рыхлого губчатого вещества, покрытого тонким слоем компактной костной ткани. Вместо надкостницы, наружный слой эпифиза формирует гиалиновый хрящ.
Если составные части кости имеют различную конструкцию и структуру, то и расти они должны по-разному. Рост трубчатой кости в длину, как уже было сказано, осуществляется за счет эпифизарной пластинки - хрящевая ткань пополняется путем новообразования клеток и их последующим окостенением. На рис. 15 отрезок А – это уже окостеневший участок хряща, отрезок В – хрящ, образовавшийся в процессе деления клеток хрящевой ткани. Рост трубчатой кости в ширину происходит за счет деления клеток надкостницы.
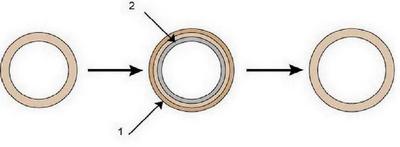
Если бы происходило лишь отложение костной ткани на наружной поверхности трубки (наружный слой 1 на рис. 17), то масса кости по мере роста увеличилась бы настолько, что животные передвигались бы с большим трудом и очень медленно. Но в то время как остеобласты наращивают костную ткань снаружи, другие клетки - остеокласты разрушают и рассасывают кость изнутри (внутренний слой 2, рис. 17), тем самым, расширяя костномозговую полость. Диаметр трубки увеличивается, но толщина костной ткани растет незначительно. Губчатое вещество эпифиза представляет собой наслоение костных перекладин (пластин) различной толщины, расположенных под разными углами друг к другу, чтобы давление и растяжение, распределялись равномерно на всю кость (рис. 15). Рост эпифизов можно сравнить с изготовлением папье-маше. Если вы возьмете мячик и станете наклеивать на его поверхность листики бумаги, располагая их равномерно по всей поверхности и обязательно под разными углами, то вы получите в результате увеличенную копию исходного объекта. Должен ли быть согласован рост диафиза и эпифизов? Обязательно! По-другому просто не может быть. Может ли рост трубчатой части повлиять на рост сустава? Вне всякого сомнения. Статистика OFA подсказывает нам, что чем больше кость растет в длину и чем меньше при этом она растет в ширину, тем более идеальный сустав получится в конечном итоге. Факторы роста, заставляющие кость расти в ширину сверх нормы для данного биологического вида, должны обязательно слегка исказить пространственную форму сустава. Это как если бы при изготовлении папье-маше мы вдруг стали наклеивать бумагу не равномерно по всей площади поверхности, а где-то больше, где-то меньше. Причастность факторов роста к формированию тазобедренного сустава помогает нам понять, почему в рамках породы одни собаки страдают дисплазией, а других минует чаша сия. Нет на свете двух собак с одинаковым геномом, а тем более, с одинаковым протеомом. Следовательно, нет двух собак, которые росли бы совершенно одинаково. Разное сочетание факторов роста и разные сочетания прочих белков, влияющих на факторы роста, дают неповторимое и оригинальное течение процесса полиферации (деления) клеток в организме. Таким образом, главное значение имеет не только общий размер и вес породы и отдельных ее представителей, но и гармоничное соотношение скорости роста трубчатых костей в длину и в ширину. В некоторых крупных породах, у тех же догов, кости растут правильно «в нужное время и в нужном месте», а в породах с другим экстерьером и другой генетикой, возможно, процессы роста не всегда совпадают, то ли по времени, то ли в пространстве. Мы можем вмешаться в процесс роста щенка и повлиять на формирование его экстерьера. К сожалению, это влияние чаще бывает негативным: недостаточное или избыточное кормление; неполноценный, несбалансированный рацион; недостаточные, избыточные или несбалансированные физические нагрузки. Но тот, кто не поленится овладеть этим великим искусством правильного выращивания щенков, сможет не только создать идеальные условия для реализации наследственной программы, но даже корректировать некоторые врожденные недостатки. Например, у щенка плоские ребра и узкая для данной породы грудная клетка, да еще один или оба родителя также имеют подобный недостаток. Казалось бы, даже не стоит пытаться что-то исправить, ведь гены, по меткому народному выражению, пальцем не раздавишь. И все же не стоит сдаваться раньше времени, активные (но строго дозированные, чтобы не навредить в чем-то другом!) физические нагрузки неизбежно приведут к увеличению объема легких. Какая бы ни была генетическая программа для роста костей скелета, но ФР из клеток легких передадут сигналы к окружающим мышцам и реберным костям: « Господа, вы должны слегка раздвинуться вширь, нам не хватает места». И ребра обязаны будут прислушаться к сигналу и скорректировать свою программу роста. Ну и напоследок еще стоит сказать, что обновление костной ткани происходит всю жизнь. У собак этот процесс особенно активен и ежегодно заменяется до 10% костной ткани. Для нормального костеобразования и рассасывания (резорбции) кости собака должна двигаться. Не полчаса в день на коротком поводке, а иметь возможность хотя бы несколько раз в неделю побегать вволю на свободе. |
| Igor A. Levakin © 2010 |